К таким «тихоокеанским» чертам шаньдиндунцев относятся значительная высота мозговой коробки (136—150 мм), альвеолярный прогнатизм и некоторая тенденция к широконосости. ЕСЛИ же учесть большую высоту лица всех шаньдиндунских черепов (69—77 мм), то наиболее близкие аналогии им встречаются только в Восточной Азии и в полярной Америке. Очень возможно, что люди, обитавшие в конце древнего каменного века на севере современного Китая, — предки позднейших популяций, на базе которых складывались восточные, а может быть, и арктические монголоиды.
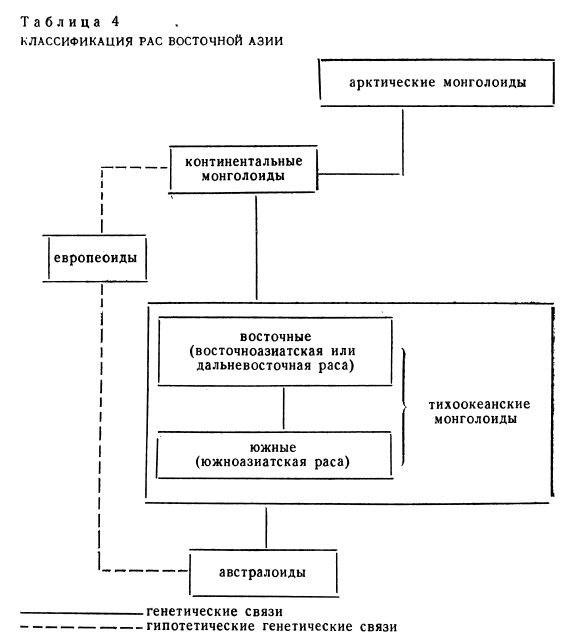
Однако сравнение шаньдиндунских черепов с люцзянским показывает, что на юге современного Китая в эпоху позднего палеолита были распространены расовые типы, отличавшиеся от северокитайских меньшими абсолютными размерами, более градильным общим обликом, несколько более узким и гораздо более низким лицом, относительной широконосостью. Таким образом, уже тогда среди тихоокеанских монголоидов наметились две группы популяций: восточная, географически и морфологически переходная к континентальным монголоидам, представленным в позднем палеолите фрагментом лобной кости из стоянки Афонтова гора II [Дебец, 1946, 73—76], и южная, связывавшая монголоидов с австралоидами (табл. 4).
Наличие известной преемственности между ранне- и поздне-палеолитическими популяциями Восточной Азии подтверждается археологическими материалами, в частности хозяйственно-культурными остатками деятельности шаньдиндунских людей. В Верхней пещере обнаружено множество разнообразных каменных орудий типа чопперов и чоппингов, костяных и роговых изделий, в том числе полированных кусков оленьих костей и рогов, один из которых напоминает «начальнические жезлы» европейского палеолита, а также различных орнаментированных предметов. Среди последних особенно интересны крупные каменные «бусины» с отверстиями, просверленные клыки оленя, лисы, дикой кошки и других хищников, костяные подвески, сверленые речные и морские раковины, доставленные сюда с морского берега за несколько десятков километров. В целом индустрия Шаньдиндуна при сохранении некоторых архаичных черт производит впечатление очень поздней, вероятно близкой к переходному периоду от палеолита к мезолиту [Pei Wen-ehung, 175—179].
Вопрос об отношении людей современного вида в целом, живших на территории Китая и Восточной Азии, к архантропам и палеоантропам того же региона очень сложен и не может считаться окончательно решенным. Сторонники гипотезы полицентризма, например Ф. Вайденрайх, К. Кун, а также почти все китайские антропологи считают, что между древнейшими, древними и современными людьми Дальнего Востока существует непосредственная генетическая преемственность. Как уже упоминалось выше, эти исследователи монголоидные расы производят непосредственно от синантропов и их среднепалеолитических потомков [Weidenreich, 1943; Coon, 1963; его же, 1965]. Среди советских ученых близкую гипотезу поддерживает В. П. Алексеев, допускающий существование в древнем казенном веке двух основных очагов сапиентации и расообразования — западного (атланто-средиземного, или афроевразийского), хронологически более раннего, и восточного (тихоокеанского или восточноазиатского), несколько более позднего [Алексеев В. П., 1967, 71—86; его же, 1969, 12—24; его же, 1970, 37— 40]. По мнению В. П. Алексеева, появление современного человека произошло в двух местах. Первое из них — Передняя Азия, возможно, с прилегающими районами; второе — междуречье Хуанхэ и Янцзы с прилегающими районами. В Передней Азии сформировались предки европеоидов и негроидов, в Китае — предки монголоидов [Алексеев В. П., 1970, 37; его же, 1974, 115—164].
Однако гипотезы о формировании Homo sapiens в нескольких (хотя бы двух) самостоятельных или почти самостоятельных очагах на основе разных подвидов архантропов и палеоантропов находятся в противоречии с открытыми еще Ч. Дарвином общими закономерностями эволюции всего органического мира под воздействием естественного отбора и не согласуются с твёрдо установленным единством всех древних и современных сапиентных человеческих популяций. В конечном счете полицентризм во всех его разновидностях приводит к телеологической или, по существу, «номогенетической» концепции эволюции человека и формирования его рас. Однако «с позиций современной биологии теория номогенеза некорректна, как понимают это определение в физике, т. е. находится в противоречии с другими, точно установленными фактами. И поэтому она должна быть отброшена» [Яденко-Хмелевский, 65].
Конкретные палеоантропологические данные также не дают достаточных оснований для вывода о специфическом родстве восточноазиатских архантропов и палеоантропов с монголоидами, особенно если учесть уже известные нам данные о всеобщем распространении лопатообразной формы резцов у древнейших и древних людей. Очевидно, правы советские антропологи Я. Я. Рогинский, В. П. Якимов, М. И. Урысон, А. А. Зубов и другие, которые стоят на точке зрения «широкого» моноцентризма, считая, что сапиентация развертывалась на обширной, но все же непрерывной территории, включающей Юго-Западную и частично Южную Азию с прилегающими регионами Северо-Восточной Африки и Юго-Восточной Европы [Рогинский, 1949, 1—156; его же, 1951, 153—204; его же, 1956, 11— 17; его же, 1970, 34—37; Урысон, 1969, 150—160; Якимов, 1967, 1—55; Зубов 1973а, 92—106].
Вместе с тем надо иметь в виду, что археологические материалы позволяют проследить преемственность хозяйственно-культурного развития населения Восточной Азии на протяжении всего древнего каменного века. Нет никаких данных, позволяющих утверждать, что архантропы и палеоантропы, жившие на территории Китая, вымерли, были истреблены позднейшими пришельцами или ушли с мест своего коренного обитания.
Таким образом, создается кажущееся противоречие между данными археологии, с одной стороны, и палеоантропологии — с другой. Это противоречие, однако, отпадает, если допустить, что сапиентация, начавшаяся на рубеже раннего и позднего палеолита в Восточном Средиземноморье и соседних регионах, постепенно захватывала все новые и новые территории, по мере расселения быстро размножавшихся подвижных сапиентных популяций и их смешения с другими группами древних людей, которые вследствие этого процесса насыщались сапиентными генами и вовлекались в общий ход формирования людей современного вида и их распространения в разных направлениях, в частности из Передней Азии через Южную и Центральную Азию на восток, вплоть до берегов Тихого океана. Очевидно, Дальний Восток на протяжении всего палеолита был периферией первобытной ойкумены, куда периодически докатывались волны переселенцев с Запада, смешивавшихся, надо думать, с аборигенами. Наиболее интенсивными эти процессы переселения были в период позднего палеолита, когда люди впервые не только заселяли новые территории Восточной Азии, но и осваивали Америку и Австралию. По свидетельству некоторых археологов, на востоке Азии нет начальных стадий позднего палеолита, а там, где элементы позднего палеолита прослеживаются, они либо непременно соединяются с мустьерскими, либо носят очень поздний облик и неотличимы от следующей ступени развития каменных орудий мезолита [Григорьев, 14—19].
В рецензии на книгу В. П. Алексеева «География человеческих рас» (1974) В. М. Массон, оценивая в целом положительно этот труд, подвергает критике гипотезу о двух очагах сапиентации и о начале образования современных рас в раннем палеолите. Согласно этой гипотезе, «может сложиться впечатление, что процесс расогенеза шел наряду и параллельно с процессом антропогенеза и в каждом из двух очагов самостоятельно формировались как разные виды: в одном — синантроп, в другом — Homo sapiens. При этом, несмотря на двух-очаговый генезис, в обоих случаях складывался идентичный биологический вид» [Массон, 1975, 121]. Неприемлемость этой точки зрения очевидна, и ссылки В. П. Алексеева на роль социальных факторов и стабилизирующего отбора [Алексеев В. П., 1974, 142] не спасают положения. Роль социальных факторов в антропогенезе и расогенезе действительно очень велика. Однако если учесть, что у человека имеется около 7 млн. генов [Эфроимсон, 42—43], то представляется очень мало вероятным возникновение под воздействием этих факторов в двух самостоятельных очагах одинакового уникального сочетания такого комплекса морфологических и физиологических особенностей, который бы позволил говорить о формировании одного и того же вида Homo sapiens в различных регионах. Стабилизирующий отбор, т. е., по определению И. И. Шмальгаузена, «отбор в пользу установившейся нормы при элиминации всех заметных уклонений от этой нормы» [Шмальгаузен, 1969, 236], может, конечно, привести к появлению в разных популяциях сходных сочетаний морфологических и физиологических признаков, но вряд ли может вызвать образование идентичных видов в двух обособленных регионах. Можно вполне согласиться с В. М. Массоном, что «более естественно принять гипотезу Я. Я. Рогинского о широком моноцентризме формирования вида Homo sapiens... С нашей точки зрения, широкая миграция вида Homo sapiens в глобальном масштабе не оставляет сомнений» [Массон, 121].
Первоначальные ареальные (расовые) различия между западными и восточными популяциями людей современного вида начали складываться, вероятно, только в самом начале позднего палеолита и касались главным образом нейтральных, неадаптивных признаков с простой генетической структурой. Различия эти обязаны своим происхождением генетико-автоматическим процессам, которые стимулировались временной, но достаточно длительной изоляцией отдельных групп неоантропов, продвигавшихся на заре человеческой истории из западных районов ойкумены в восточные и ассимилировавших разные группы неандертальцев. Сформировавшиеся позднее (не ранее конца позднего палеолита) австралоидные и монголоидные расы тихоокеанского бассейна унаследовали указанные ареальные различия позднепалеолитических предков и, в свою очередь, передали их своим потомкам, у которых они сохранились, по крайней мере частично, до наших дней [Чебоксаров, Чебоксарова, 88—163; Зубов, 1973, 20—27; Чебоксаров, 1973, 3—19]. «Восточные» (монголоидные и, вероятно, австралоидные) группы популяций отличались от «западных» (европеоидных и негроидных) сохранением большей доли резцов лопатообразной формы, большим процентом круговых узоров на подушечках пальцев, специфическим распределением генов по некоторым системам групп крови (например, почти 100%-ной концентрацией резус-положительности или высокой частотой особого фактора Даффи) и многими другими одонтологическими, дерматоглифическими и серологическими особенностями [Воронов, 85—89; Cheboxarov, Zoubov, 1-18].
Можно предполагать, что уже в позднем палеолите «восточные» группы популяций не были однородны в расовом отношении. Популяции северной части современного Китая (к северу от Циньлина) принадлежали скорее всего к восточноазиатской (дальневосточной) расе, которая составляла северную ветвь тихоокеанских монголоидов. Именно к этой расе, как мы видели, должны быть отнесены позднепалеолитические черепа из Шаньдиндуна. Учитывая, что во все последующие эпохи различные типы этой расы были наиболее характерны для населения Тибета и всего бассейна Хуанхэ, надо, по-видимому, искать истоки этносов китайско-тибетской языковой семьи среди популяций восточноазиатской расы. К северу от восточных монголоидов в пределах Внутренней Монголии, частично также Синьцзяна и Маньчжурии, жили в позднем палеолите, вероятно, различные популяции континентальных монголоидов, ареал которых простирался в пределы современной МНР и Сибири (к ним относится фрагмент детского черепа из стоянки Афонтова гора II в Красноярске) [Дебец, 1946, 73—76].
В южной половине современного Китая в позднем палеолите обитали, вероятно, различные популяции южной ветви тихоокеанских монголоидов, переходных к австралоидам. Промежуточные монголоидно-австралоидные морфологические черты, как мы знаем, очень характерны для люцзянского человека. На базе южномонголоидных подуляций, входящих в южноазиатскую расу, сложились в более поздние исторические эпохи тайские (чжуан-дунские), мяо-яоские, аустроазиатские (монкхмерские) и аустронезийские этносы, область формирования которых была расположена, по мнению многих исследователей, в Южном Китае и Северном Индокитае. Еще южнее — в Индонезии и, возможно, на Филиппинах — в позднем палеолите жили, надо думать, австралоидные популяции, представленные черепами из Ваджака, Ниа и Аитапе. Возможно, что популяции эти говорили на языках, близких к папуасским и северохальмахерским [Решетов, Чебоксаров, 89—114].